
搜索结果: 1-15 共查到“生物学 编码区”相关记录26条 . 查询时间(0.185 秒)
中国农业科学院植物保护研究所科研人员开发内源基因非编码区定向进化新技术
内源基因 非编码区 定向进化 新技术
2024/4/22
近日,中国农业科学院植物保护研究所作物病原生物功能基因组研究创新团队开发了核酸酶LbCas12a介导的内源基因非编码区定向进化技术。相关研究成果发表在《植物通讯 (Plant Communications) 》。
中国科学院生物物理研究所专利:一种针对东亚人群全基因组范围内的非编码区的SNPs的DNA芯片
中国科学院生物物理研究所专利:一种针对欧洲人群全基因组范围内的非编码区的SNPs的DNA芯片
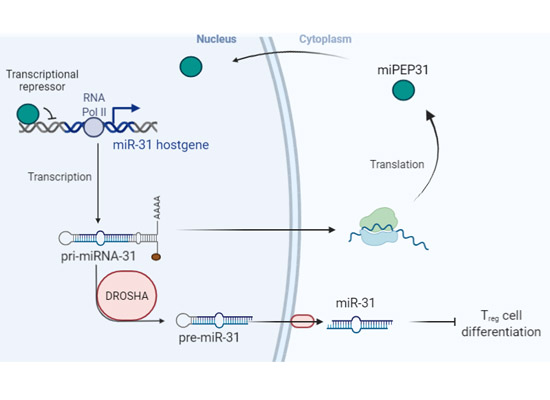
上海交通大学医学院附属第一人民医院临床研究院王宏林团队在EMBO Reports再次发表基因组非编码区编码多肽治疗自身免疫性疾病的创新性成果(图)
王宏林团队 自身免疫性疾病 全身性的疾病 核糖体印记
2023/1/13
自身免疫性疾病通常被认为是既是器官特异性又是全身性的疾病,这些疾病的发生主要是由于免疫系统的故障误攻击了自身的细胞和组织。大约8%-10%的人口受到自身免疫性疾病的影响,这种疾病会导致严重的损伤、高死亡率和高医疗费用。尽管现有的治疗方法(如小分子和抗体药物)有助于减缓疾病进展及减少残疾的发生,但它们会导致严重的副作用,且不能逆转疾病的表现。因此,寻找新的创新治疗方法和药物分子是目前转化医学研究的首...
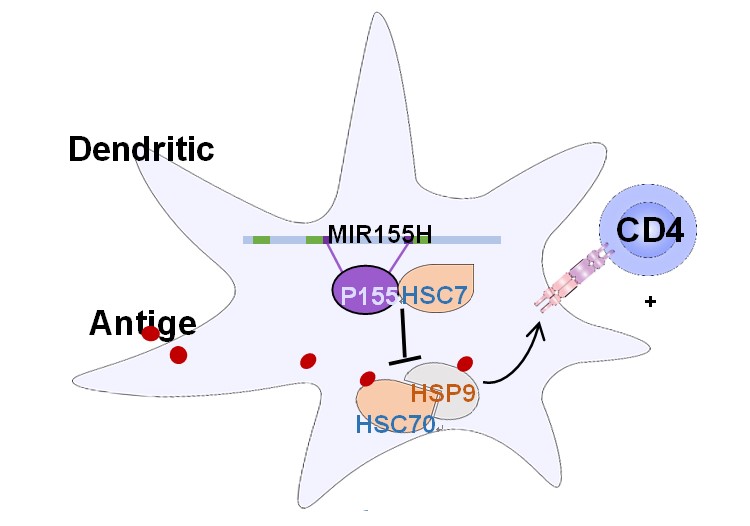
上海交通大学医学院/上海市免疫学研究所王宏林课题组在Science Advances发表论文,原创性发现人基因组“暗物质“(非编码区编码的多肽)具备治疗自身免疫性疾病的潜能(图)
银屑病 王宏林课题组 暗物质 人树突状细胞
2023/4/13
2020年5月20日,上海交通大学医学院/上海市免疫学研究所王宏林研究员团队在Science Advances发表了题为“A Micropeptide Encoded by LncRNA MIR155HG Suppresses Autoimmune Inflammation via Modulating Antigen Presentation”的研究论文,报道了课题组原创性发现人基因组非编码区编...

四川农业大学动物科技学院一种克隆山羊TNNC2基因编码区全序列的方法(图)
川农业大学 动物科技学院 克隆山羊 方法
2019/5/29
专利名称:一种克隆山羊TNNC2基因编码区全序列的方法;所有发明人:陈浩林、徐刚毅、汪代华、徐洪刚

如何解读与复杂疾病有关的遗传变异,尤其是非编码区变异,是疾病遗传学领域的一个巨大挑战。现有的方法在评估非编码区致病性变异方面存在着假阳性率过高、敏感度不够等问题。为了解决这一难题,中国科学院北京生命科学研究院赵方庆团队首次提出一种全新的基于人群等位基因频率谱的监督集成算法(PAFA),以实现对复杂疾病和性状相关的遗传变异进行打分识别及功能性评估。7月11日,该团队的最新研究成果以Prioritiz...

西安交大生命学院杨铁林教授课题组以骨质疏松症疾病为切入点,揭开了基因组“暗物质”非编码区1p36.12基因座的调控秘密:通过整合基因组、转录组、表观组和三维基因组的系统生物学分析,确立了1p36.12区潜在关键功能性调控变异――SNP-rs6426749。结合数据和功能实验,研究发现,该SNP位于一个增强子上,rs6426749-G碱基通过结合转录因子TFAP2A,激活增强子活性,进而通过染色质成...
高原鼢鼠神经型一氧化氮合酶基因编码区序列克隆与分析
高原鼢鼠 神经型一氧化氮合酶 基因克隆 序列分析
2016/7/18
高原鼢鼠是青藏高原特有的地下鼠,受到高原低氧以及洞穴低氧的双重低氧环境压力。经RNA 提取、RT-PCR、亚克隆与测序,本研究获得高原鼢鼠神经型一氧化氮合酶(nNOS)的编码区序列,并对其分子特征进行了分析。结果显示:高原鼢鼠nNOS 基因编码区(CDS)全长4 290 bp,编码1 429 个氨基酸残基;CDS 与大鼠、小鼠、兔、狗、人的同源性分别为90% 、89% 、87% 、87% 、89%...
猪瘟病毒编码区基因选择压力及重组分析
猪瘟病毒 编码区全基因组 适应性进化 重组
2014/1/7
为了更全面地探究选择压力和基因重组对猪瘟病毒基因组全编码区进化的影响,本研究通过生物信息学软件包PAML 4.5中的codeml程序和RDP 3.44中的6种重组检测算法,对采集时间1994~2010年,涵盖3个基因型中的6个基因亚型,遍布11个国家的50株猪瘟病毒(CSFV)基因组全编码区进行了选择压力及重组分析. 结果显示编码区基因所编码的全部12个蛋白质的选择压力测度参数ω均低于1,表明CS...
藏羚羊PGC-1α基因编码区的克隆与分析
cDNA克隆 PGC-1α 能量代谢 藏羚羊
2012/3/22
以藏羚羊(Pantholops hodgsonii)及同海拔分布的藏系绵羊(Tibetan Sheep)的心肌组织为材料,提取总RNA,利用逆转录聚合酶链反应(RT-PCR)技术扩增出过氧化物酶体增生物激活受体γ辅激活因子-1α(PGC-1α)的基因编码区cDNA片段,与载体连接构建重组质粒,经转化、扩增培养、鉴定后测序。利用生物信息学方法分析显示,藏羚羊和藏系绵羊的PGC-1α基因编码区长度均为...
藏羚羊组蛋白去乙酰化酶1基因编码区的克隆与序列分析
藏羚羊 组蛋白去乙酰化酶1 基因克隆 低氧适应
2016/7/20
为探讨藏羚羊的低氧适应机制与高原低氧环境的相关性,采用RT-PCR 技术,首次从藏羚羊心肌组织总DNA 中克隆出组蛋白去乙酰化酶1 (Histone deacetylase1,HDAC1)基因的编码区序列。该序列全长为1 449 bp,编码482 个氨基酸,预测其蛋白质分子量约为55 kDa。序列分析表明,藏羚羊HDAC1 基因的编码区序列与其它哺乳动物相似性超过90% ,其中与牛的相似性最高为9...
近平滑念珠菌ERG 11基因编码区的克隆及生物信息学分析
近平滑念珠菌 ERG11基因 克隆测序
2012/8/10
目的 克隆、测序近平滑念珠菌ERG11基因的编码区序列并进行生物信息学分析。方法 运用生物信息学的方法 ,通过与白念珠菌ERG11基因碱基序列同源性比对,在近平滑念珠菌基因组(www.sanger.ac.uk/sequencing/Candida/pa-rapsilosis/)中寻找可能的ERG11基因序列(CpERG11),并据此序列设计引物,经PCR扩增近平滑念珠菌标准株(ATCC22019)...
Yeast基因组编码区特征参数的研究
Yeast基因组(Yeast genome) 编码区(Gene coding region) 碱基成分偏移(Bias count of base component) 正相关(Direct proportion)
2008/3/5
以碱基成分偏移量D值?眼1?演为基本参数定义参数d,以d为Yeast编码区的特征参数,对Yeast的第1、2、3类ORF(open reading frame)进行了统计,得到d的特征参数区间。并且,以此区间为标准对Yeast的6类ORF,以及5′帽、3′尾、内含子、组分随机序列等非编码序列进行了检验。结果表明,用d作编码区的特征参数是可行的,它可以很好地区分编码序列和非编码序列。另外,又讨论了参...